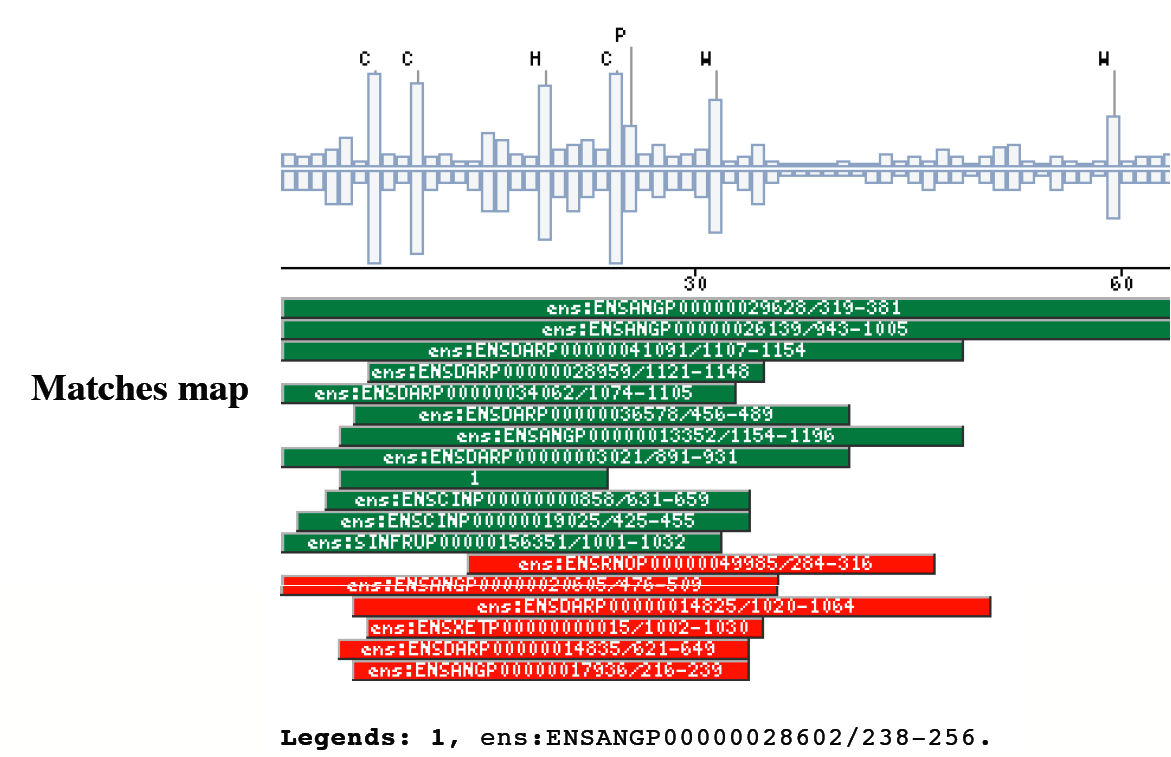
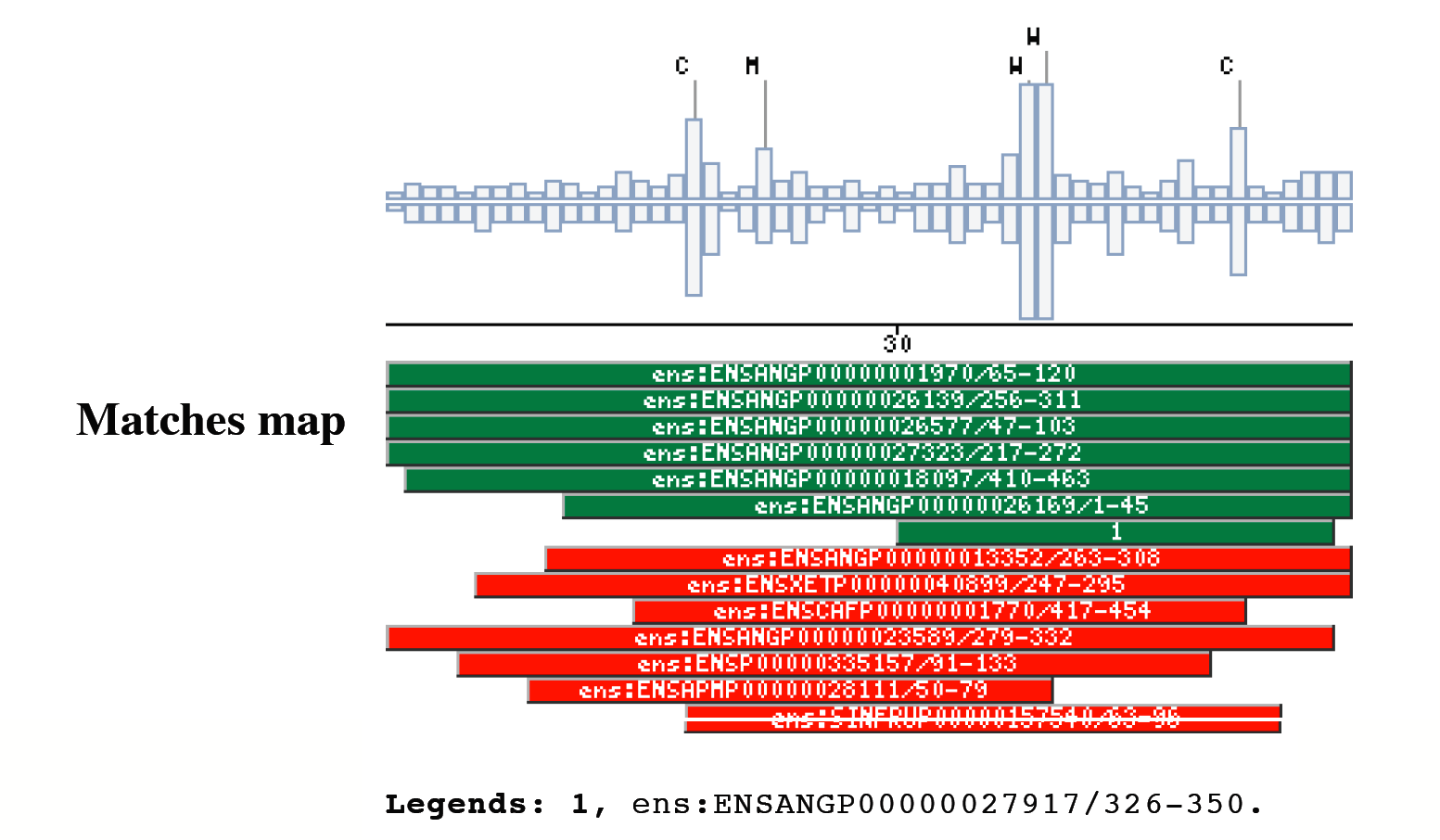
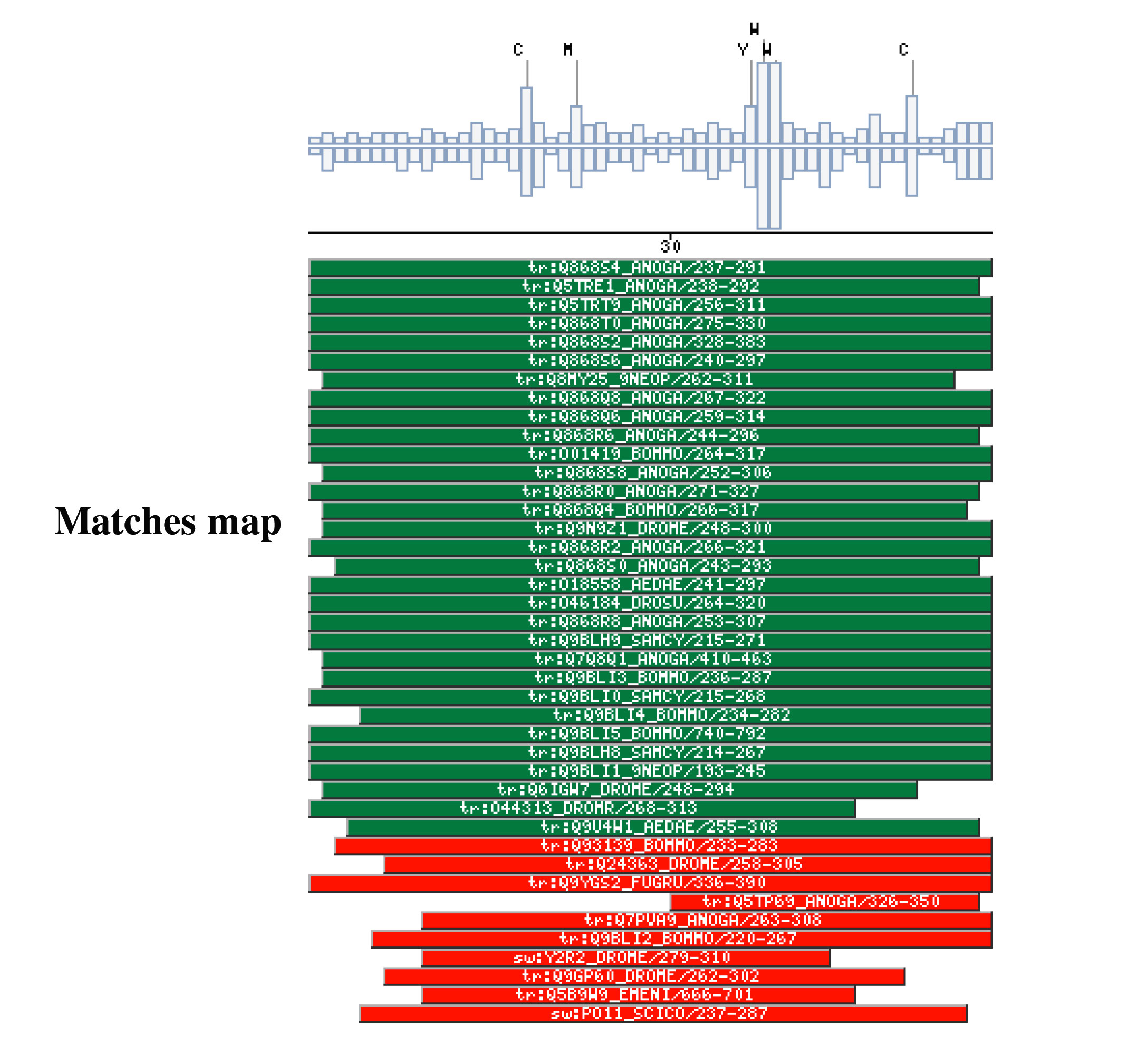
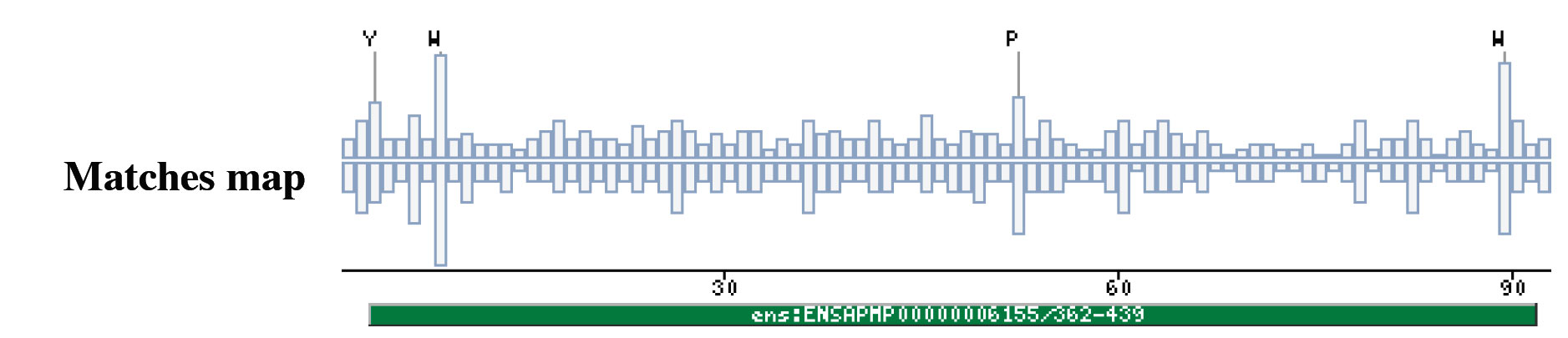
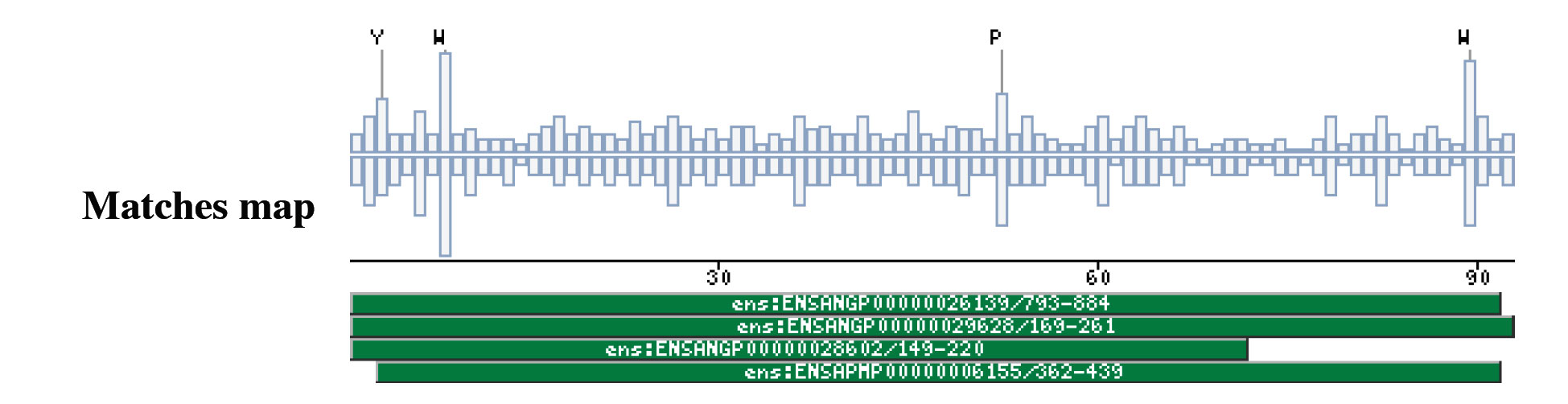
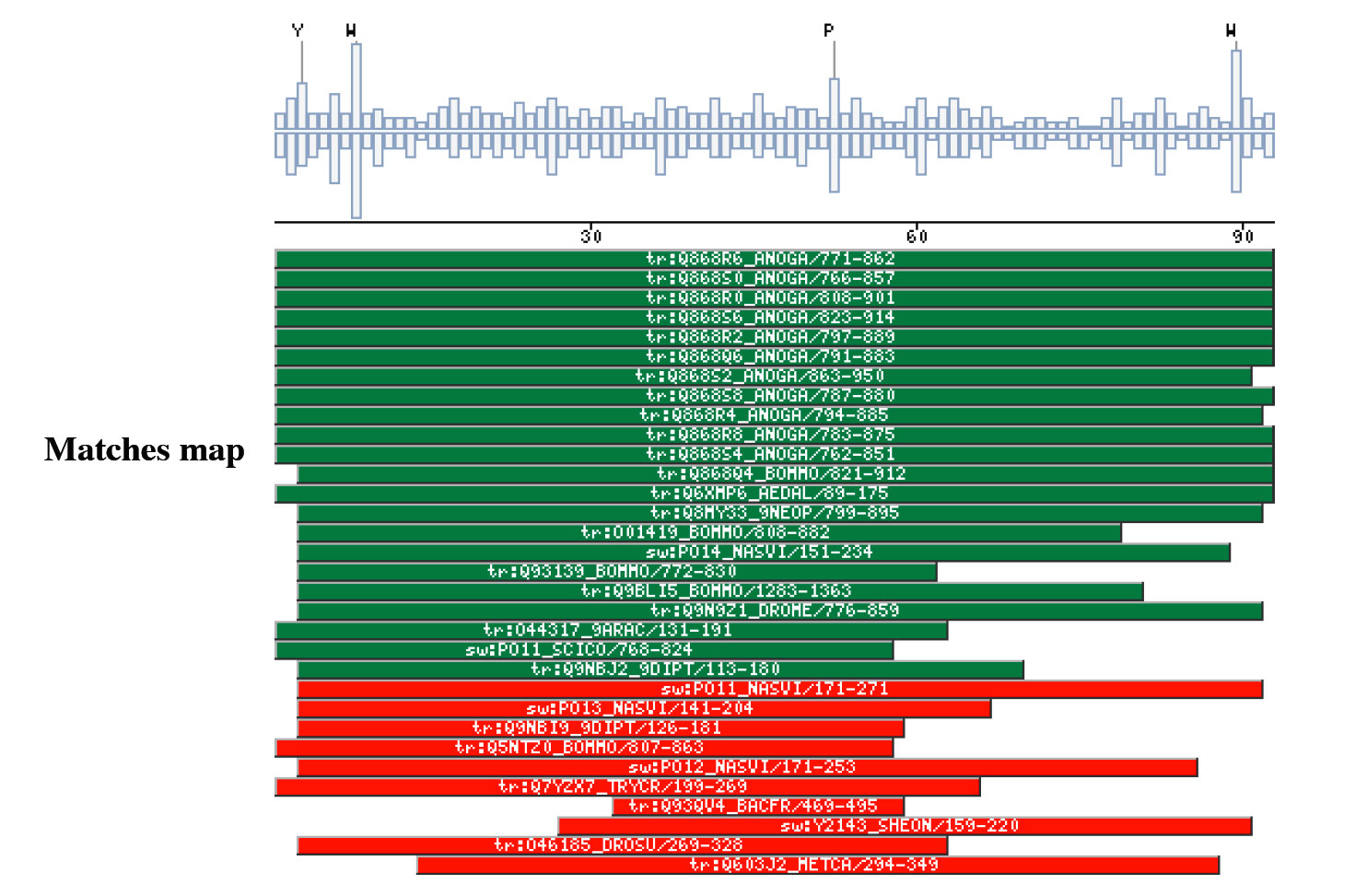
|
 |
| |
|
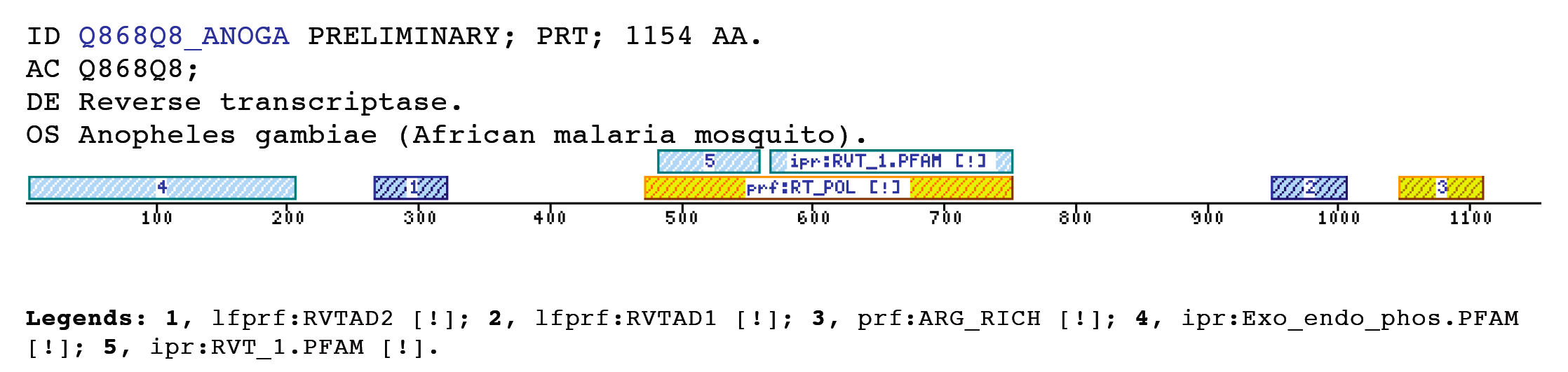
The insect non-LTR reverse transcriptase caseTransposable elements make up a substantial proportion of the total DNA in most, if not all eukaryotic genomes (Wright and Finnegan 2001). Those elements fall in two classes: the retrotransposons that transpose by a process involving reverse transcription and the transposons that move by an excision-insertion mechanism. Retrotransposons are of two types, the best known resemble retroviral proviruses in having long terminal repeats (LTRs) and open reading frames equivalent to gag, pol and in some cases, env genes of retroviruses (Wilhelm and Wilhelm 2001). Retrotransposons of the second type, the LINEs or non-LTR elements, have no terminal repeats. These elements, originally discovered in mammalian genomes, have been detected in a wide range of species from protozoa to fungi, plants and animals, which implies that are known in almost all kind of living organism, generally have two open reading frames. There is little, if any, sequence similarity between those proteins encoded by the first of those open reading frames, although they may all be nucleic-acid-binding proteins, that are able to form ribonucleoprotein particles during transposition (Kazazian and Goodier 2002).The proteins encoded by the second open reading frame of these elements, however, are related in sequence to reverse transcriptases, and elements of this type has been shown to transpose by a process involving reverse transcription. In fact, transposition of Non-LTR retrotransposons is similar to that of the retrovirus like elements (Finnegan 1997). It has been suggest that integration takes place at sites where chromosomal DNA has been nicked by the host encoded products as might be expected to occur during DNA repair or recombination, of course this can not account for the integration of the subset of non-LTR retrotransposons that insert at specific sequences (Dawson, Hartswood et al. 1997). This subset includes the elements found only at specific sites within the miniexon genes of some trypanosomes and ribosomal RNA genes of insects (Gabriel, Yen et al. 1990). The reverse transcriptase enzyme catalyse polymerisation is an RNA-directed DNA-polymerase. Occurs in a variety of mobile elements, including retrotransposons, retroviruses, group II introns, bacterial msDNAs, hepadnaviruses, and caulimoviruses. The retroviral reverse transcriptase is synthetized as part of the POL polyprotein that contains; an aspartyl protease, a reverse transcriptase, RNase H and integrase. POL polyprotein undergoes specific enzymatic cleavage to yield the mature proteins (Inouye and Inouye 1995). The two found domains related with this kind of proteins are insect specific. The location of these domains is as follows: RVTAD1: is always located between the domains: pfam Exo_endo_phosp (PF03372) and pfam:RVT (PF00078). RVTAD2: C-ter of pfam: RVT (PF00078). The domain is located within the nucleic acid binding endonuclease region of PO14_NASVI. RVTAD3: In PO14_NASVI this domain overlaps with the reverse transcriptase region, but in the other cases (Q868S0_ANOGA, PO11_SCICO) is located toward the C-terminus of this domain. The reverse transcriptase region is described by the profile RT_POL (PS50878), which describes the catalytic domain of this kind of proteins, and the domain Pfam: RVT_1 (PF00078). Examples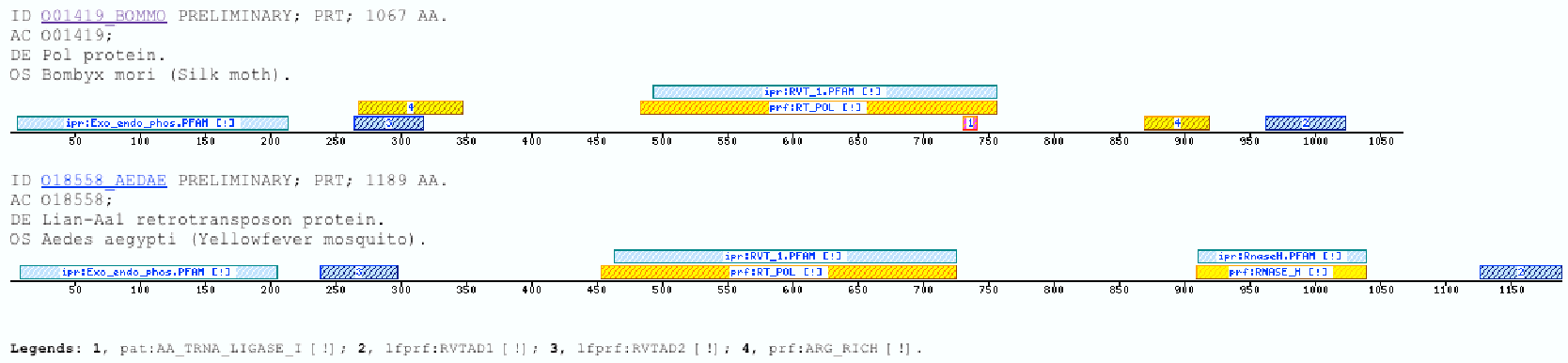

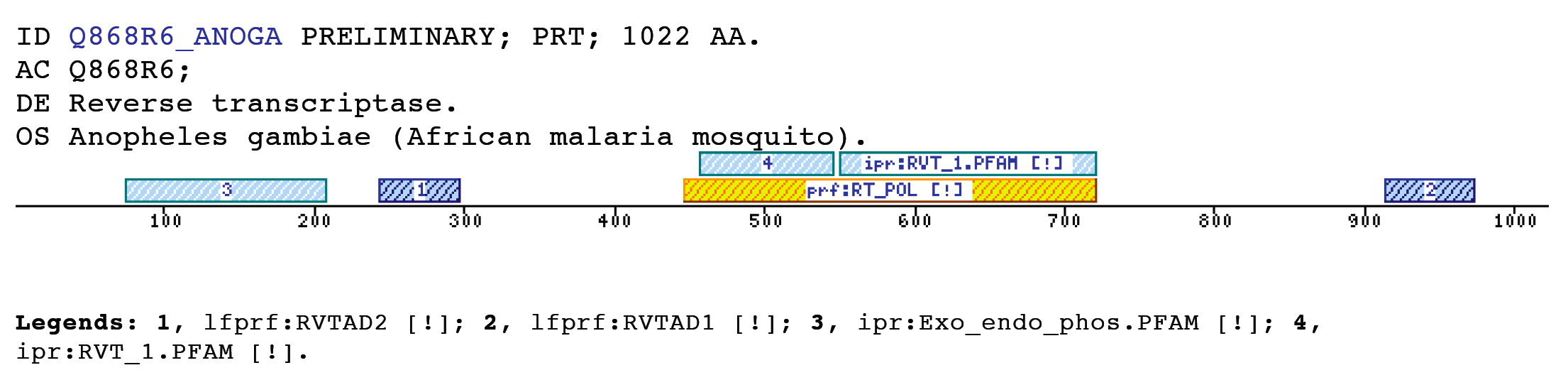
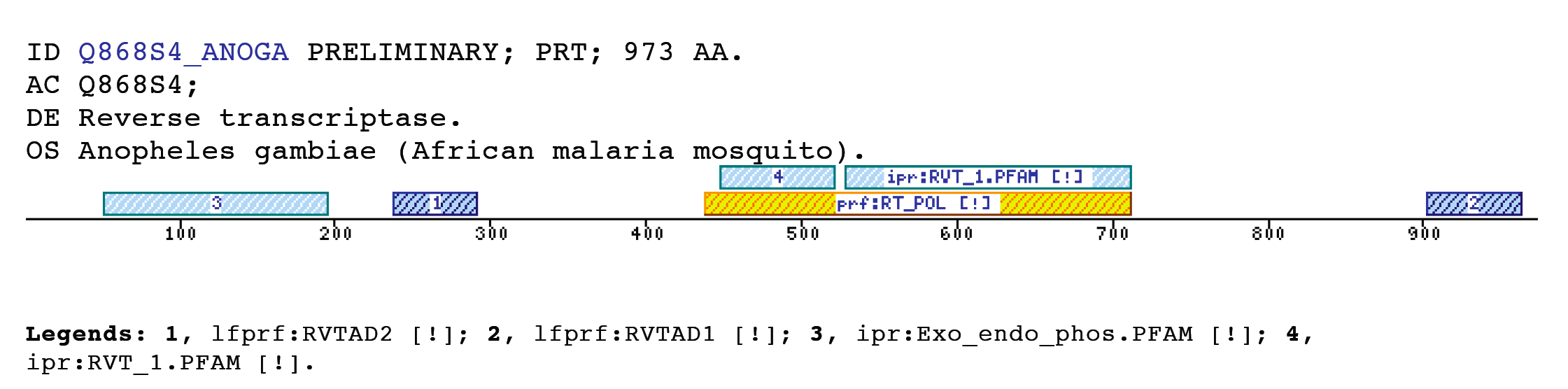
Taxonomic DistributionRVTAD 1Insects: Papilio xuthus, Nasonia vitripennis, Anopheles gambiae. Plants: Arabidopsis thaliana, Oryza sativa, Zea mays, Brassica campestris. Protozoa: Entamoeba histolytica. RVTAD 2 and 3 Insects: Sciara coprophila, Anopheles gambiae, Papilio xuthus, Aedes aegypti.
REFERENCESDawson, A., E. Hartswood, et al. (1997). "A LINE-like transposable element in Drosophila, the I factor, encodes a protein with properties similar to those of retroviral nucleocapsids." Embo J 16(14): 4448-55. Finnegan, D. J. (1997). "Transposable elements: how non-LTR retrotransposons do it." Curr Biol 7(4): R245-8. Gabriel, A., T. J. Yen, et al. (1990). "A rapidly rearranging retrotransposon within the miniexon gene locus of Crithidia fasciculata." Mol Cell Biol 10(2): 615-24. Inouye, S. and M. Inouye (1995). "Structure, function, and evolution of bacterial reverse transcriptase." Virus Genes 11(2-3): 81-94. Kazazian, H. H., Jr. and J. L. Goodier (2002). "LINE drive. retrotransposition and genome instability." Cell 110(3): 277-80. Wilhelm, M. and F. X. Wilhelm (2001). "Reverse transcription of retroviruses and LTR retrotransposons." Cell Mol Life Sci 58(9): 1246-62. Wright, S. and D. Finnegan (2001). "Genome evolution: sex and the transposable element." Curr Biol 11(8): R296-9. |